
Я полагал, что понимание принципов систематики (здесь речь о макросистематике, т. е. о систематике надвидовых таксонов) является для биолога само собой разумеющимся. Но недавно в беседе с одним коллегой я обнаружил, что кто-то из нас — или я, или он — эти принципы не понимает. Тогда я решил посмотреть русскоязычную литературу по систематике. Результаты меня не обрадовали. Если бы было секретное задание написать про систематику так, чтобы, прочтя, читатели подумали: какой автор умный, а я ничего не понял, увы… — то следовало бы писать именно так. Нужно было свалить в кучу немногие актуальные концепции и многие давно отправленные на помойку, замучить ненужными подробностями и бесчисленными цитатами. Вот характерный пример такого стиля. Открыв статью «Кладистика» в русскоязычной «Википедии», читаем: «Кладистика относится к числу трех ведущих таксономических школ, доминирующих в современной биологической систематике; ей противостоят фенетика, основанная на количественной оценке так называемого общего сходства, и эволюционная таксономия, которая, подобно кладистике, при построении системы опирается на эволюционную близость (т. е. общность происхождения), однако не требует строгого соответствия системы и филогении…» Вот это мило! А география случайно не представлена двумя школами? Одна школа — обычная география, а другая — школа сторонников плоской формы Земли? Я пришел к выводу, что мне следует высказаться на страницах ТрВ-Наука, тем более что истина столь проста, что может уложиться в небольшую заметку.
Давайте договоримся о следующем: современная систематика основана на филогенетике, т. е. на общности происхождения, и только на ней. Работая над такой систематикой, мы работаем над выяснением истории развития жизни. Такие знания не менее важны для удовлетворения присущего нашему виду любопытства, чем прояснение истории самого человечества. Знание филогенетической систематики помогает решить и множество других вопросов, как теоретических, так и практических. Все остальные подходы к систематике, какими бы заумными словами они ни прикрывались, недалеко ушли от объединения жирафа и журавля по признаку длинношеестости или поскольку оба на «Ж». Есть немного биологов, которые этого не понимают, и много больше биологов, которые отказываются это принять по причинам, я думаю, психологического свойства. Спорить тут бессмысленно; тем, кто не согласен, что систематика должна отражать только эволюционную историю жизни, дальше читать не имеет смысла.
Теперь перейдем к собственно кладистике. Кладистика — метод построения систематики.
Она предлагает несколько принципов, которые кажутся очевидными, когда их знаешь. Однако сформулировать эти принципы смогли только в 1950 году. Сделал это немецкий энтомолог Вилли Хенниг. (Человеколюбие не позволяет мне рекомендовать читателям первоисточник. Хенниг сочинил кладистику как побочный продукт, а занимался он таксономией мух. Поскольку я работаю с теми же семействами, с которыми работал Хенниг, то его тексты мне знакомы не понаслышке. Они написаны на безумно тяжелом немецком языке.)
Итак, принципы кладистики. Первый принцип уже назван выше: систематика отражает эволюционную историю, и ничего, кроме нее. А как же нам заглянуть в прошлое, чтобы эту эволюционную историю восстановить? Рассмотрим какую-нибудь реальную кладограмму — на них можно смотреть вечно, как на огонь и волны. Вот высшие обезьяны; человек изображен выше всех, как и подобает венцу творения. Старые систематики часто рассуждали в парадигме «примитивный — прогрессивный»; такой подход нередко давал хорошие результаты, но страдал глубокой произвольностью выбора. Вот человекообразные обезьяны дружно утратили хвосты — это прогрессивно? Лиса так не думает, я тоже сомневаюсь. Хенниг предложил изучать филогению в парадигме «сначала — потом»: она и целям соответствует, и понимается всеми одинаково. Человека можно поместить хоть вверху, хоть внизу, сути это не изменит. Суть в том, что первыми (из нарисованных тут) разделились 31 млн лет назад таксоны мартышкообразных и человекообразных, это сестринские таксоны самого высокого ранга. А последними 6 млн лет назад разделились ветви шимпанзе и человека, это сестринские таксоны самого низкого (тут) ранга.
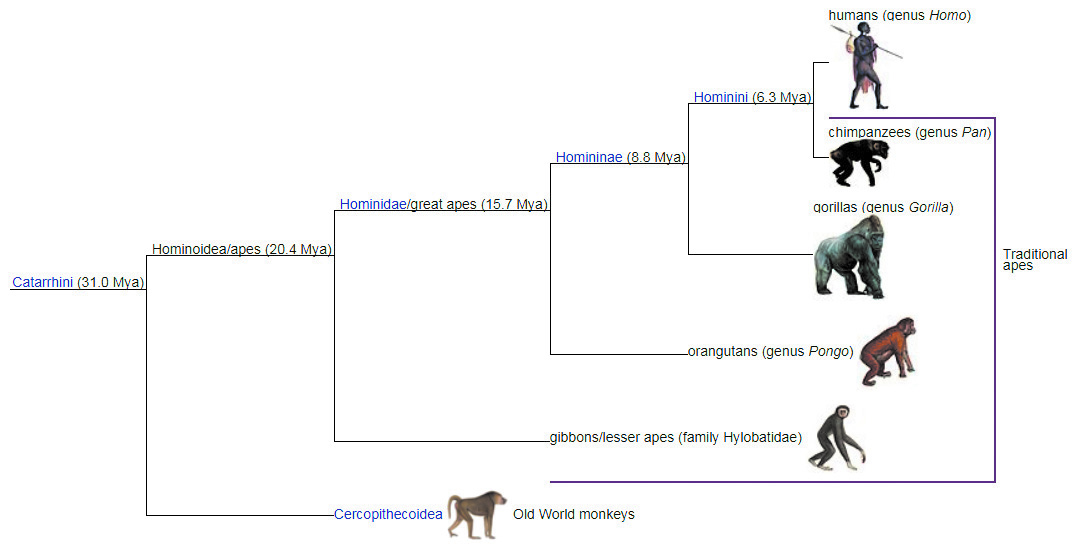
Если вернуться к хвостам, то даты ветвлений указывают, что начальным (плезиоморфным, по Хеннигу) состоянием было «обезьяны с хвостами», а потом отделилась ветка с новым (апоморфным) состоянием «хвост утрачен». А если бы не было данных молекулярных часов, как их не было во времена Хеннига? Мы всё равно могли бы резонно предположить, что хвостатость плезиоморфна (изначальна), а бесхвостость апоморфна. Почему не наоборот? Потому что большинство обезьян хвостаты, большинство млекопитающих тоже. Потому что утратить хвост много легче, чем приобрести его. Но мы не рассуждаем в логике: человек — венец творения, поэтому, если у него нет хвоста, бесхвостость прогрессивна. Еще раз: термины Хеннига обретают смысл, только будучи наложенными на шкалу времени. Нормальные млекопитающие используют ноги для ходьбы, это плезиоморфное состояние. А летучие мыши переднюю конечность укоротили, пальцы удлинили, растянули между ними перепонку и теперь летают — это синапоморфия (общая апоморфия) летучих мышей. Но по отношению к рыбам уже четвероногость — апоморфия, а рыбья безногость — плезиоморфия. Но безногость змей снова апоморфия по отношению к предшествовавшему состоянию четвероногости их предков.
Наконец, третий принцип кладистики, вытекающий из первого. Очевидно, что систематика будет отражать филогенез в том и только в том случае, когда соблюдено нехитрое правило: корректный таксон включает всех потомков некой ветки и никого из потомков других веток. Такой таксон называется монофилетическим (также голофилетическим в русскоязычной литературе). В нашем случае монофилетическими будут, например, таксоны: Pan, включающий обыкновенного и карликового шимпанзе; Pan + Homo, включающий человека и шимпанзе; Hominoidea, включающий всех бесхвостых обезьян.
Немонофилетические таксоны по недостатку (= взяли не всех потомков ветви) называются парафилетическими, немонофилетические таксоны по избытку (= взяли кого-то с чужой ветки) называются полифилетическими. Немонофилетический таксон как по заказу изображен на нашем рисунке и обозначен Traditional apes.
Как видите, ничего особо нового в кладистике нет, просто последовательное и строгое соответствие систематики и филогенеза. Термины «апоморфия — плезиоморфия» не употреблялись, но, по сути, классические биологи старались рассуждать именно так. До всего, до чего можно было додуматься, классики додумались, упомянутые выше таксоны человекообразных обезьян или летучих мышей ни у кого сомнений не вызывали.
А потом, в 1977 году (через год после смерти Хеннига), Карл Вёзе и Джордж Фокс сделали замечательное открытие. Авторы сравнили последовательности нуклеотидов рибосомальной РНК и обнаружили, что прокариоты делятся на две клады. Это собственно бактерии и группа, преимущественно специализирующаяся на экстремальных местообитаниях, — археи. Вскоре выяснилось, что выявленное по рРНК разделение подтверждается и массой других отличий; сегодня этот факт является несомненно установленным. Кстати, наш (эукариотов) ядерный геном унаследован в основном от архей, а геномы митохондрий и пластид имеют бактериальное происхождение. Открыть новое надцарство организмов — это круто, конечно, но главное не в этом. Во-первых, обнаружено самое древнее ветвление филогенетического дерева жизни, точнее, самое древнее из таких, потомки обеих ветвей которого дожили до наших дней. Ниже этой точки ствол дерева по отношению к рецентным (т. е. невымершим) организмам прямой и исследованию филогенетическими методами недоступен. Во-вторых, за миллиарды лет эволюции изначальные последовательности могли бы неузнаваемо измениться. На наше счастье, некоторая часть генов оказалась удивительно консервативной и содержит информацию о далеком прошлом. В-третьих, суперконсервативные участки генома позволили обнаружить архей, а супервариабельные — помогают подтвердить или опровергнуть отцовство, например. Из этого следует, что при правильном выборе последовательностей всё филогенетическое дерево может быть реконструировано молекулярными методами, вопрос только во времени и силах, которые для этого потребуются. Чем современная биология и занимается: программы обсчитывают последовательности, а биологи интерпретируют результаты.
Что дает мне нахальную уверенность в том, что мое понимание принципов систематики верное, а моего коллеги — нет? А вот что. Программе не объяснишь, что существуют «три ведущие таксономические школы», ей надо какую-нибудь одну задать. Догадываетесь какую? Не ту, которая «опирается… однако не требует…», а ту, которая сформулировала имеющую смысл и решаемую задачу: выбрать из возможных филогенетических деревьев то, последовательность ветвлений которого наиболее вероятна.
Программа работает примерно так: взяли k последовательностей, после выравнивания в них нашлось n вариабельных локусов. Проверим последовательно гипотезы, что самой ранней апоморфией была замена в локусе 1, в локусе 2, в локусе n. Перебрав варианты, найдем самое простое дерево, которое объясняет имеющиеся замены. (На практике обычно используют более экономичные методы, например метод максимального правдоподобия, но сути это не меняет.) Когда и если сторонники «альтернативных школ» смогут предложить логику рассуждений, из которой следует иной способ обсчета, тогда будет смысл вести с ними дискуссию.
* * *
Давайте проверим, удалось ли мне понятно изложить суть систематики на одной полосе.
1. Как сделать, чтобы таксон Traditional apes на рис. 1 стал монофилетическим?
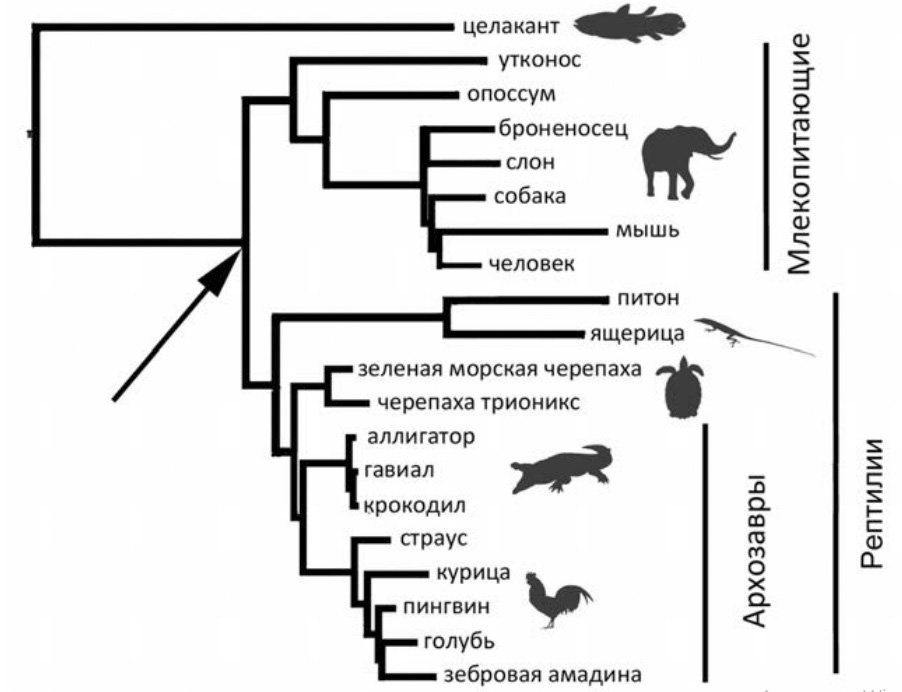
2. На рис. 2 — кладограмма из книги «Евангелие от LUCA» Максима Винарского. Пояснения автора таковы: «Одной из первых „жертв“ [кладистики] пали ни в чем не повинные птицы. Если взглянуть на родословное дерево пресмыкающихся, то мы увидим, что птицы не образуют самостоятельного эволюционного ствола: эта группа животных помещается в самой середине генеалогического древа пресмыкающихся, причем ближе к крокодилам, чем к ящерицам или черепахам…»
Во всем ли вы согласны с автором?
Никита Вихрев,
канд. биол. наук
Что значит искусственно объединенными? Общий корень у нептичьих завропсид есть? Есть. Те физиологические, анатомические и экологические признаки, которые объединяют их в группу «рептилий», плезиоморфны? Да. Поэтому не объединение «искусственное», а исключение птиц, с точки зрения кладистики, «искусственное». Но учитывая совершенно другую, апоморфную, физиологию, анатомию и экологию птиц, исключение вполне оправданное, если мы занимаемся современными, а не ископаемыми животными.
У птиц нет «совершенно другой апоморфии, физиологии, анатомии и экологии». Кроме птиц, были прочие теплокровные рептилии, которые вымерли. Для исключения птиц из числа рептилий нет никаких оснований ни по генотипу, ни по фенотипу. Класс птиц должен быть упразднен, или крокодилы, чешуйчатые и черепахи выделены в отдельные классы.
Уважаемые читатели, честно говоря я ожидал более решительных возражений. Д-р Гельфанд справедливо возразил мне, что кто именно «пал жертвой» — вопрос спорный.
Кстати, если уж совсем по кладистике, то идеальная систематика будет: ящерицезмеи и черепахи+крокодилоптицы. Но у меня много знакомых орнитологов, я не хочу чтобы меня придушили.
Как человек далёкий от проблем таксономии, не понял в чём острота вопроса. Если бы «народная» таксономия называла птиц и змей одним словом, а черепахи так не назывались, то были бы проблемы. А по картинке имеем монофилетические таксоны: птицы, крокодилы, черепахи, змеи с ящерицами. Всех их вместе можно обозвать «рептилоиды».
Вероятно, проблема в слове «рептилии», они же «пресмыкающиеся», под которыми птицы не подразумеваются.
Ну так и назвали бы как-нибудь более нейтрально. Термин не должен создавать проблемы. Птиц вмести с крокодилами и динозаврами обозвали архозаврами и никто вроде бы не возражает.
Тут нужно сделать ряд уточнений:
Как раз в молекулярке максимальная парсимония уже почти не используется. А так всё верно.
За пределами молекулярки тем более :)
Нет, вообще-то, в палеонтологии и морфологической филогенетике максимальная парсимония доминирует.
Простите, я совсем не специалист во всем этом, но по-любительски давно интересуюсь систематикой животных. Вот такой вопрос, наверное он и к птицам имеет отношение, но мне легче его проиллюстрировать на китобегемотах. Кладистика помещает их, если я не ошибаюсь, в один таксон, но ветви китов и бегемотов разделились много десятков миллионов лет назад, причем их общих предок вроде не был похож ни на тех, ни на других, киты, к примеру, независимо долго эволюционировали.. то есть, несмотря на наличие общего предка, ветви давно разделились, независимо долго проэволюционировали и породили много видов.. Интуитивно кажется, что такая ситуация сильно отлична от той, когда за тот же промежуток времени общий предок смог породить только немного похожих видов, тогда как кладистика это не учитывает, и в том, и в другом случае животных помещают в таксон одного ранга. Ну вот и вопрос — нельзя ли как-то модифицировать
современный подход к систематике, чтобы учеть подобные разницы?
Ваш вопрос касается не кладистики (т.е. кладистического анализа как метода), а применения линнеевских рангов к кладистической классификации. Еще сам Хенниг пришел к выводу о том, что никак не получится объективно применить ранги к кладам, почему и отказался от них, использовав в последние годы безранговую систему (т.е. никаких типов, классов и отрядов, только пронумерованные клады). Причин тут великое множество, начиная с того, что точек ветвления внутри клады может быть неспопоставимо больше всех мыслимых рангов, какие приставки не добавляй, и заканчивая тем, что какой «объективный» критерий не возьми (саму последовательность ветвления, как вы описали, или возраст клады, или её субъективное морфологическое своеобразие), с другим критерием из этого списка возникнет противоречие. На практике чаще используется последний критерий, но можно смело сказать, что присвоение конкретным кладам конкретных рангов — дело вкусовщины и практичности (полезности системы для потребителей-несистематиков).
Из этой ситуации пытались выйти, разработав новый кодекс номенклатуры, уходящий от линнеевской системы рангов, PhyloCode, но, как метко выразился Эндрю Брауэр, он получился мертворожденным. Поэтому нужно просто принять субъективность рангов и не париться по этому поводу. Содержание — оно в том, кого именно помещают в кладу, а не в том, какой ранг ей присваивают.
Большое спасибо за ответ! В принципе я согласен не париться, но ведь действительно, для потребителей-несистематиков, а тем более, вообще,
для неспециалистов, старая система как-то привычнее.. Нельзя ли ее усовершенствовать, присваивая отдельным рангам какие-нибудь индексы,
учитывающие, например, их возраст и приблизительное количество видов?
Всего, разумеется, не учтешь, но, для потребителей возможно будет полезно.. Хотя, может это и бред..
Реальные потребители, в первую очередь, там, где это практически важно, обычно следуют традиции. Ни одного орнитолога вы не убедите, что птицы — это не класс. Все они признают птиц классом и считают ранги от него «вниз» по филогении. То же с термитами. Диктиоптерологи могут сколько угодно выделять их в своих ранговых классификациях как семейство или надсемейство тараканов, изоптерологи всё равно продолжат считать их отрядом, т.к. система группы за 100 лет разработана очень подробно, и закрывать глаза на практически важные клады, потому что по строго кладистической и при этом ранговой системе для них не хватило рангов, просто глупо.
Хотя некоторые мои коллеги считают иначе, моё личное мнение таково, что ранги никогда не получится сделать объективными так, чтобы все были довольны — в линеевской системе с её смешным количеством ступеней иерархии реальному биоразнообразию всегда будет тесно.
ОК, большое спасибо за ваше мнение!
> Что дает мне нахальную уверенность в том, что мое понимание принципов систематики верное, а моего коллеги — нет? А вот что. Программе не объяснишь, что существуют «три ведущие таксономические школы», ей надо какую-нибудь одну задать.
Такой хороший вопрос и такой плохой ответ.