Никогда позвоночные не завоевали бы сушу, если бы не смогли на ней размножаться. Самки рыб и амфибий, как правило, мечут икру в воду, а самцы оплодотворяют кладку. Но на суше яйцеклетки нужно защищать от высыхания, они спрятаны в глубинах материнского тела, и сперму приходится доставлять туда. Внутреннее оплодотворение потребовало наружных половых органов, и они появились. Их происхождение исследовано гораздо хуже, чем появление конечностей. Лапы развились из плавников, а гениталии откуда?
В ноябре 1997 года в журнале Nature появилась статья под названием «О пальцах и пенисах» (Of fingers, toes and penises). Ее авторы, швейцарские и американские исследователи под руководством Дени Дюбуля (Denis Duboule) из Женевского университета, обнаружили, что развитие пальцев и формирование наружных половых органов мышиного эмбриона контролируют одни и те же гены Hoxd и Hoxa. Эти гены активно работают в зачатках передних и задних конечностей и мочеполовой системы, а если они не функциональны, животное получается и без пальцев, и без гениталий.
Естественно, возникло предположение, что между лапами и наружными половыми органами существует эволюционная связь, и биологи к этой идее неоднократно возвращались. Недавно американские и французские ученые под руководством Клиффорда Табина (Clifford J. Tabin), возглавляющего отдел генетики Гарвардской медицинской школы, и Жерома Гроса (Jérôme Gros) из Института Пастера исследовали развитие и эволюционное происхождение наружных половых органов самцов наземных позвоночных. Оказалось, что, хотя в некоторых случаях связь между задними лапами и гениталиями существует, дело все-таки не в ногах, а в клоаке эмбриона.
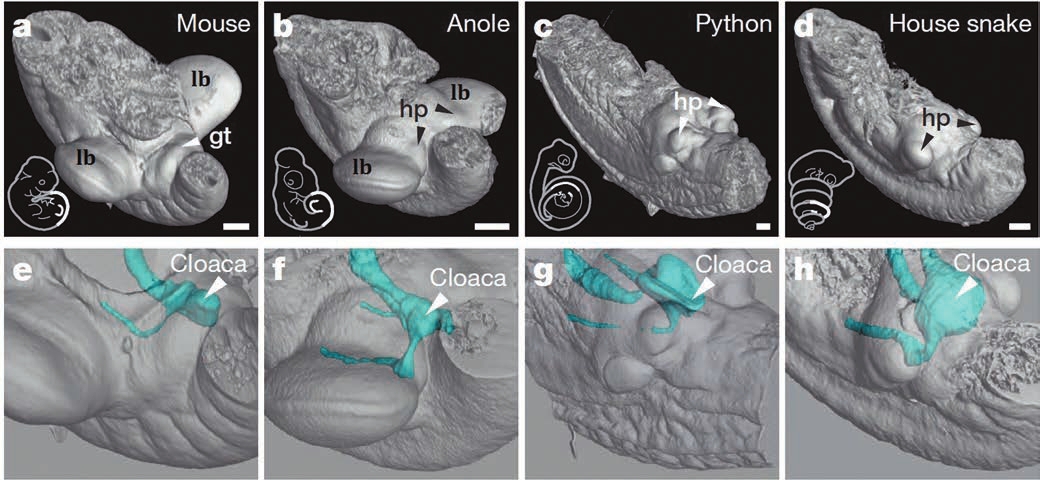
Исследователи начали с того, что определили взаимное расположение зачатков конечностей и половых бугорков, из которых развиваются пенис млекопитающих или гемипенисы (парные половые органы) рептилий, в эмбрионах мышей, ящериц семейства анолисовых (Anolis), питона и домовой змеи. У змей конечностей нет, но их зачатки в ходе эмбрионального развития образуются. Как показала микрокомпьютерная томография, у чешуйчатых половые бугорки возникают вплотную с зачатками задних конечностей. А поскольку зачатков два, то и половых бугорков тоже два, а у взрослых самцов рептилий два гемипениса (при спаривании они пользуются одним).
Развитием гениталий управляют сигнальные молекулы, которые синтезируют клетки эмбриональной клоаки (структуры, из которой затем формируются мочевыводящий тракт и кишечник). Она у рептилий также находится рядом с конечностями. У мышиного эмбриона клоака смещена к заднему концу тела (хвостовой почке), туда же сдвинулся половой бугорок, он один и с зачатками лап не связан. Из него впоследствии образуется один пенис.
Действительно ли различие в положении развивающихся органов отражает разницу в их происхождении? Исследователи это проверили, вводя в разные участки эмбрионов мыши, цыпленка и анолисовой ящерицы вектор с геном зеленого флуоресцентного белка и наблюдая, в какие клетки переместится метка. Оказалось, что у ящериц конечности и гемипенисы развиваются из одной ткани. Эти органы светятся зеленым, если метку вводили в целóм эмбриона. Когда метили хвостовую почку, ни конечности, ни половой бугорок не окрашивались.
Иными словами, наружные половые органы рептилий действительно растут почти оттуда же, откуда и ноги. У мышей конечности формируются из той же популяции клеток, что и у ящериц, а половой бугорок — из другой ткани, которую образуют клетки, расположенные рядом с хвостовой почкой. У цыплят ситуация промежуточная. Инъекция в целом куриного эмбриона метит и конечности, и половой бугорок, в хвостовую почку — только бугорок.
Обнаруженную разницу подтвердило и сравнение генов, работающих в зачатках конечностей и гениталий на разных этапах эмбрионального развития. У ящериц наборы мРНК, которые синтезируются в этих тканях, сходны и на ранних, и на поздних стадиях, что неудивительно, учитывая общее происхождение зачатков. Лишь в поздней стадии, когда начинается формирование гениталий, в них включаются гены, специфические для этого органа. У мышей и на ранних, и на поздних стадиях ткани конечностей и пениса четко различаются по набору активных генов, хотя и небольшое сходство между ними есть.
Ученые предположили, что место образования гениталий зависит от положения эмбриональной клоаки. Она синтезирует сигнальные молекулы, действующие на близлежащие клетки, из которых и образуются наружные половые органы. У рептилий клоака находится между зачатками задних ног, у птиц она начала перемещаться к хвостовой почке, а у млекопитающих окончательно «сползла». Поэтому у рептилий два гемипениса между ногами, а у млекопитающих один, ближе к хвосту. У примитивных млекопитающих, таких как утконос, пенис один, но раздвоен.
Эту гипотезу исследователи проверили, пересадив помеченные зеленым флуоресцентным белком клетки эмбриональной клоаки цыпленка или перепела в зачатки задних конечностей другого эмбриона. Спустя 1–2 дня на них появлялись дополнительные бугорки. Они образовывались из клеток эмбриона-реципиента, так что клетки пересаженной клоаки не сами делились, а влияли на деление окружающих клеток. Активность генов в этих дополнительных бугорках изменилась и приобрела характерные для гениталий особенности. Когда клетки пересадили в ткань хвостового бугорка, дополнительные зачатки и экспрессия генитальных маркеров возникли там. Разные типы эмбриональных клеток одинаково реагируют на сигнальные молекулы клоаки, благодаря чему в них реализуется одна генетическая программа. Наружные половые органы млекопитающих и рептилий негомологичны, хотя и выполняют сходные функции, но в память о былом родстве гениталий и конечностей они еще используют некоторые общие гены, регулирующие их развитие.
Tschopp P., Sherratt E., Sanger T.J., Groner A.C., Aspiras A.C., Hu J.K., Pourquié O., Gros J., Tabin C. J. A relative shift in cloacal location repositions external genitalia in amniote evolution // Nature. 2014. doi: 10.1038/nature13819